2.5 第二の飛躍:アルゴリズムの発動
・アルゴリズムとはなにか:自然現象としてのアルゴリズム
生命誕生に関するいろいろなモデルを見てきたが、ここで、一つ試したいことがある。それは、計算科学などで使われる「アルゴリズム」の概念を拡張し、それを自然現象の因果的な連鎖の順序系列と見なし、その視点から「進化」のアルゴリズムを分析することである。それによって、生命や進化のしくみをより深く理解できるのではないかと思うのだ。
「アルゴリズム」という単語を聞いたことがないかもしれないので、アルゴリズムの意味から説明したい。アルゴリズムは計算科学の言葉で、簡単に言えば「計算手順」のことである。難しそうに聞こえるかもしれないが、日常生活で誰もが使っている。
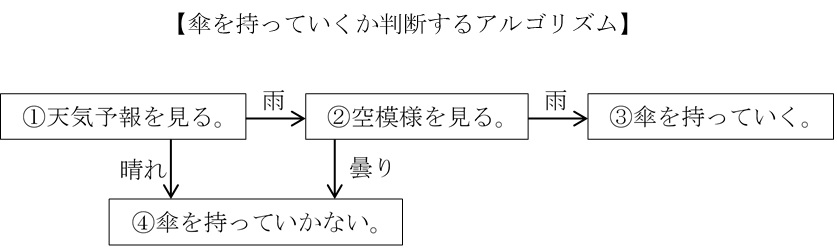
例えば、朝、家を出るときに傘を持っていくべきかどうか判断するとしよう。この問いの答えを求めるとき、何らかの手順にしたがって答えを出すだろう。私の場合、まず天気予報を見て、雨の予報の場合、玄関の外に出て空模様を見る。そして、すでに雨が降っていれば傘を持っていくが、雲だけなら持っていかない。このような方法で判断するわけだが、この方法は以下の手順(アルゴリズム)として表せる。
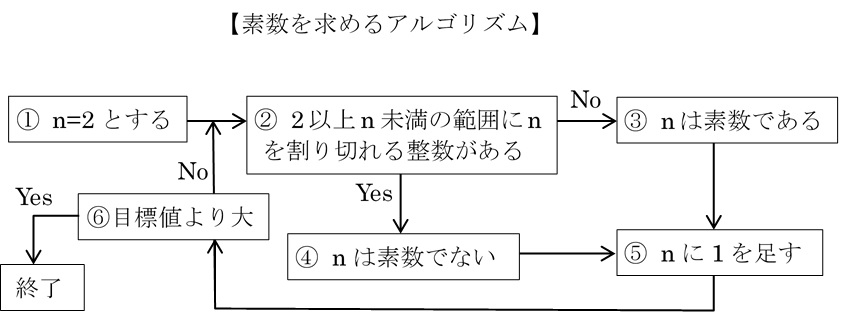
多くのコンピューター・プログラムは、アルゴリズムを骨格として作られている。例えば、素数を求めるプログラムを作ろうとしたとき、さまざまな導出方法が思いつくが、次のアルゴリズムに従えば、目標とする値以下の範囲ですべての素数を求めることができる。
このようにアルゴリズムとは計算手順のことである。
ここで、アルゴリズムの概念を拡張し、「自然現象の因果的な連鎖の順序系列」のことだと考えてみよう。この「自然アルゴリズム」は、個々の自然現象の性質でも、物体の性質でもなく、状況や条件が持っている性質である。そう考えると、連鎖的に起きる自然現象を一つのアルゴリズムとして分析することができる。
例えば、「雪崩」を自然アルゴリズムとして表すと次のようになる。
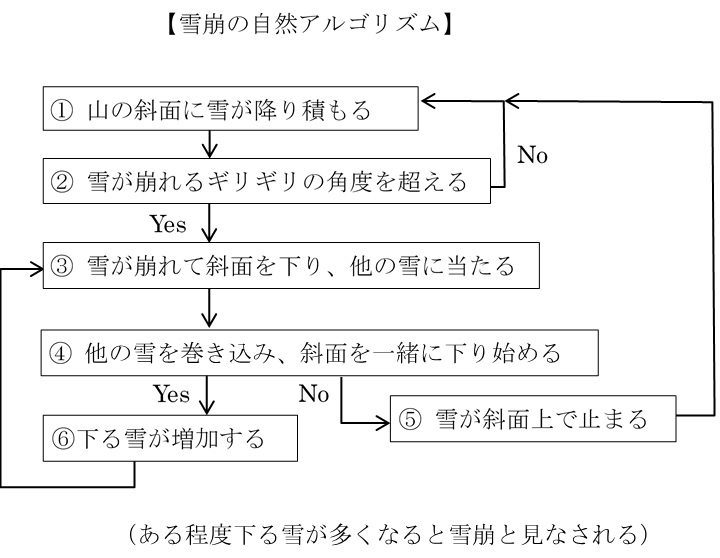
こうした自然現象の連鎖である「自然アルゴリズム」もネゲントロピーを消費しながら起きる。最初にあった位置エネルギーや運動エネルギーがすべて熱化(散逸)してネゲントロピーを使い切れば、現象の連鎖は止まる。
他の例としてエルニーニョ現象も挙げられる。この場合、「南米沖での海水温上昇」→「熱帯域の海水温低下」→「太平洋高気圧の弱化」→「日本の冷夏」のようなアルゴリズムと見ることができる。このようにさまざまな自然現象の連鎖をアルゴリズムとして見ることは可能であり、それによって諸現象を統一的に見ることができるようになる。ただし、この見方は新しいものではない。複雑系科学の中では当然のように用いられてきた方法である。
先に述べたイングランドの「散逸適応」のモデルについても、同様にアルゴリズムとして次のように表すことができる。
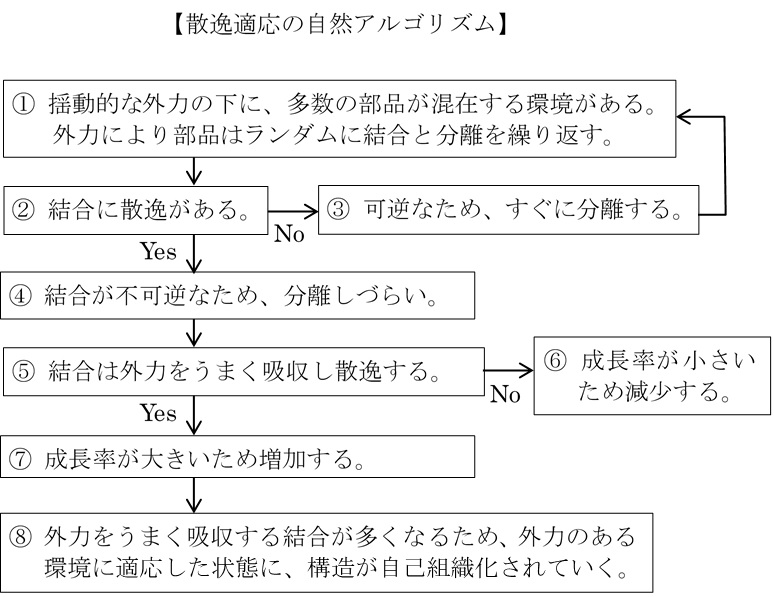
複雑系科学においてアルゴリズムとして扱われてきた現象の代表例が「進化」と「学習」である。進化は「遺伝的アルゴリズム」(遺伝現象を模倣したプログラム)を用いて研究されてきた。遺伝的アルゴリズムを用いれば、進化と類似の現象を計算機シミュレーションによって再現でき、「もしも世界が〇〇だったら」という人工的な諸条件の中での進化を試してみることができる。
同様に、学習についても「ニューラルネットワーク」というヒトや動物の学習過程を模倣したプログラムを使って研究されてきた。進化や学習のアルゴリズムを統一的な視点から分析した研究もある。(例えば文献[**]。)
・最適化アルゴリズムの発動条件
進化や学習のアルゴリズムは、素数のアルゴリズムと比べて大きく異なる点がある。それは、アルゴリズムの中に偶然の要素(ランダムネス)が入っている点である。素数を求めるアルゴリズムのように、出発点から一つの決まった道筋だけを進んでいき、必ず決まった答えにたどり着くアルゴリズムを、ここでは「決定論的なアルゴリズム」と呼ぼう。また、アルゴリズムが偶然の要素を含み、同じ出発点から始めても、さまざまな経路を進んで、異なる答えにたどり着くアルゴリズムを「非決定論的なアルゴリズム」と呼ぶことにする。
アルゴリズムに偶然の要素(非決定論)を入れて、うまく道筋を組むと、環境や条件に最も適した解を見出すことができる。そのようなアルゴリズムを「最適化アルゴリズム」という。そこで見出されるのは、正しい一つの解ではなく、いくつかの最適な解である。
ここで、「最適」とは次のような意味である。例えば、「2+X=5を満たすXはいくつか」と問われれば、その解は一つ(X=3)に決まる。つまり、「正しい解」が存在する。しかし、いくつも解の候補があり、どれが正しいか検証するのにとてつもない時間がかかるという問題(NP問題という)もある。このとき、「正しい解」を見つけるのが非常に困難なため、いくつも解の候補を考え、そこからより適していると思われる解を見出す。その解が「最適解」である。「最適」という言葉を使っているが、ベストな解(正しい解)ではなくベターな解(探した範囲でより良い解)という意味である。
例えば、一週間の時間割を作るとする。A先生は月曜日学校に来ない、B先生は1時間目の授業ができないなど、さまざまな制約を考慮した上で、先生方が最も喜ぶ時間割を作りたいとする。こうした時間割編成の問題はNP問題になることが知られており、時間割の候補数は膨大になる 。そのすべての候補を比較して「正しい答え」を見出すのは不可能である。それよりも、有限な時間でより良い答え(最良とは限らない)を選び出す方が賢い方法だと言える。それが「最適化」という方法である。
最適化の方法は生命も使っている。ある環境に住む場合、最良の方法を選択するのは難しい。何万通りも選べるさまざまな方法の中で、その方法が最良かは確かめようがない。だから、生命の個体は「やってみる」という方法を採る。そして、うまく生き残られれば、それはその環境において良い方法だったということになる。このような現象を(環境への)「適応」という。
最適化(あるいは、最適化アルゴリズム)を自然現象と見なしたとき、進化や学習は最適化という現象の一種であり、「適応」は進化という最適化過程の一部だと見ることができる。
「最適化」という自然現象はどのような条件が成り立つときに発現するだろうか。その発生条件について考えてみよう。まず、進化がどのような自然現象であるのか思い出そう。進化の要点だけ取り出すと、次のようにまとめられる。(単純化しすぎだと進化生物学者に叱られるかもしれないが。)
(Evo1) 遺伝子:個体は分子配列(遺伝子)の中に情報(遺伝情報)を保持している。同種の遺伝子型を持つ個体の集まりを種という。遺伝情報は個体の外形など(表現型)を決めており、環境への適応度に違いを生み出す。
(Evo2) 自己複製・突然変異:個体(遺伝子)は自己複製を行い、数を増やす。自己複製の際、遺伝子情報の一部にランダムな変化が起きる。
(Evo3) 自然選択:個体は環境と相互作用する。適応度の高い個体(遺伝子)は生き残り、適応度の低い個体は淘汰される。
上記(Evo1)〜(Evo3)によって遺伝子情報が少しずつ変化して、種の個体の形質(体形など)が少しずつ変化していく。それが「進化」だと見なすことができる。
学習についても考えよう。こちらも専門家に叱られるかもしれないが、要点を大胆に切り取ってまとめてみる。
(Learn1) 神経回路網:神経回路網はその構造の中に情報(内部情報)を蓄えられる。内部情報は外界からの入力に対する反応の違いを生み出す。
(Learn2) 回路変化:神経回路網は神経回路の一部をつなぎ変えることができる。
(Learn3) 選択的強弱:神経回路網は外部入力に反応する。適切な反応の場合、その神経系を強化し、不適切な場合、その神経系を弱化させる。
上記(Learn1)〜(Learn3)によって内部情報が少しずつ変化し、同じ入力に対する反応がより良いものへと変化する。これが「学習」だと考えられる。
進化や学習においてそれぞれ上の3要素が重要だと思われるが、情報や計算という視点で見たとき、それらはどのような役割を果たしているだろうか。
(Opt1) 情報保持:システムが安定的な内部構造を持つとき、その構造の中に内部情報を保持することができる。
(Opt2) ランダムネス:システムは弱いランダムネス(転写ミスや配線つなぎ変えなど)があることにより内部構造を徐々に変化させることができる。
(Opt3) 取捨選択:内部構造が含む情報の違いは、外部に対するシステムの応答の違いを生み出す。応答が適しているとき、その変化は強化され、適していないとき、弱化される。
これらの素過程が連鎖すると、なぜ進化や学習などの「最適化」(最適化アルゴリズム)という現象が発動するのか考えてみよう。まず、情報保持についてだが、システムが安定した構造(DNA配列や神経回路網など)を持つと、そこに何らかの情報を記録できる。ここで言う「情報」とは、外部への応答を作る際に参照される構造のことである。安定した構造が情報(の容器)になるということだ。その構造の状態は情報空間中の一点で表され、情報空間は配列(DNA配列や神経回路網の配線)の要素の数だけ次元を持つ。構造の安定性は情報空間の安定性を意味する。
次に、ランダムネスについて考えると、システムに弱いランダムネスがあれば、安定した構造に少しずつ変化が加わる。それは情報を内蔵する構造の分布がいろいろな方向に少しずつ拡大することであり、情報空間中で一点に集中していたサンプル群が徐々に拡散することを意味する。これは、情報空間での探索範囲の拡大に相当する。
最後に、取捨選択についてだが、これが最も重要だと言えよう。唯一、外部と関係する過程ともいえる。システムは外部とやり取りし、外部からの入力に何らかの応答を返す。このとき、内部構造が持つ情報を外部への応答に反映させる。応答が適切な場合、対応する内部構造が強化され、不適切だと弱化される。それによって情報空間でのサンプル群の分布がより適した点へと収束する。つまり、状態が最適化される。
このように上記(Opt1)-(Opt3)の過程があるだけで、最適化という自然現象が起きる。どのように最適化が起きるのかイメージを示したのが、下に示した図2-13である。
本来、情報空間は内部構造の自由度の数だけ次元を持つが、この図では情報空間を1次元(横軸)で表した。また、最適化で得られる解は一つとは限らない。サンプルが生き残る場所は複数の場所(AとB)である。それらのうち最も多くサンプルが生き残っている場所(B)を最適解と見なす。
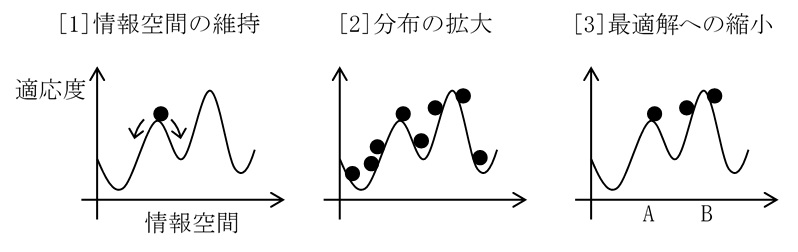
図2_13. 最適化:情報空間での最適解(A,B)への収束。縦軸は適応度を示し、曲線は適応度の地形を表す。
この最適化のしくみをアルゴリズムとして表すと次のようになる。
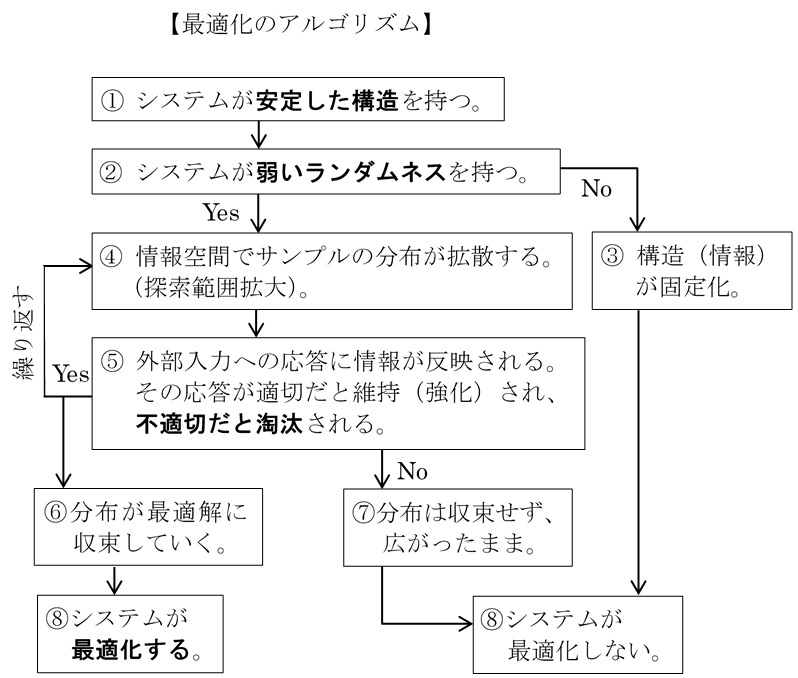
このように3つの条件がそろうと、最適化というアルゴリズムが発動し、システムは自ら環境に適した構造へと変化させていく。そう考えると、進化も学習も最適化アルゴリズムという種類の自然現象だと見なすことができる。振り返ってみれば、イングランドの散逸適応の過程も、[1]安定した構造、[2]ランダムネス、[3]淘汰の3つの要素が入っていた。これも最適化の一種だと見なせよう。
物理学から生物学、情報科学などさまざまな分野の諸現象を広く見渡すと、同様の現象がいくつも見つかる。例えば、中垣俊之が研究する「粘菌の知性」[**]も田中博の言う「バクテリアの脳」[**,p.119](粘菌やバクテリアは単細胞生物だから知能などあるはずないが)最適化という現象の一種だと見なせば、それらの賢さを理解できる。また、情報科学で良く知られた「シミュレーテド・アニーリング」という最適化の手法は、温度パラメーターと呼ばれる量で指定される(熱的な)ランダムネスを導入した最適化の手法で、これも最適化という自然現象の要点だけ取り出して模倣したアルゴリズムだと考えられる。
最適化という自然現象はいたるところで起きている。最適化(=最適化アルゴリズム)を自然現象と認めれば、生物の進化も脳の学習も単細胞生物の知性もすべて「最適化」という同種の現象であることに気づく。実際、「散逸適応」は分子の部品レベルでの最適化であるし、「生命の進化」は分子レベルでの最適化であり、「バクテリアの脳」は細胞レベルでの最適化、「粘菌の知性」は細胞集団レベルでの最適化だと見なすことができる。そして、「学習」は神経回路網レベルでの最適化である。
言いすぎだと叱られるかもしれないが、個体レベル・社会レベルでの最適化も見出すことができる。それは科学や芸術などの文化である。すでにそういう視点での議論はされていて、文化を「進化」という現象の一つだと見たとき、文化の中のそれぞれの要素に「ミーム(文化遺伝子)」という名前が付けられている。文化の要素とは、科学や芸術で世代を超えて受け継がれてく要素のことである。例えば、科学で言えば理論や法則、モデルなどであり、芸術で言えば技法や道具、個性などである。以下に示すように文化(ミーム)にも最適化の3要素がある。このように最適化アルゴリズムは大きさの階層を超えて成り立つ現象だと見ることができる。
(Mem1) 文化遺伝子(ミーム):社会は各個体の脳や記録媒体(石、紙、CDなど)の中に情報(文化遺伝子)を保持できる。
(Mem2) 自己複製と変化:文化遺伝子は個体間の伝承や記録媒体の複製によって情報が広く社会(多数の個体)に伝わる。その際、伝承ミスや意図的な改変(科学理論や芸術手法の改良)によって少しずつ情報が変化する。
(Mem3) 選択:文化遺伝子は社会の中で個体と相互作用する。より多くの評価を得た文化遺伝子は生き残り、評価の低い文化遺伝子は淘汰される。
このように見ると、文化も最適化アルゴリズムの一種だと見なせる 。
・条件は偶然、発生は必然
進化や学習は「最適化アルゴリズム」という自然現象であるという見方をここで強調するのは、それによって生命の誕生に対する偶然性と必然性の理解がさらに深まるからである。上で見たように最適化アルゴリズムという自然現象は[1]〜[3]の条件を満たしさえすれば、簡単に発生する。その意味で、生命の誕生(進化の始まり)は必然である。しかし、それらの条件を満たすことはそれほど簡単ではない。その意味で生命の誕生は偶然である。
この考え方を用いれば、地球誕生後、比較的早い時期に生命が誕生したという事実と地球外生命がまだ見つかっていないという事実の両方をうまく理解できる。地球誕生後、比較的早い時期に生命が誕生したのは、(条件がそろっていれば)生命の誕生が起きやすい現象だからである。それは、水に落としたインクの滴が水の中に広がっていくように自然に起きる。イングランドが提案した「散逸適応」のようなイメージである。
また、いまだに地球外で生命が見つかっていないという事実は、生命の誕生がなかなか起きないことを示唆している。これは最適化としての進化がなかなか起きないということではなく、最適化が起きる3つの条件が宇宙の中では簡単にそろわないということだろう。
安定した構造、弱いランダムネス、自然淘汰の3つは意外とそろいにくい。そもそも固体のように構造が安定化しすぎていれば変化が起きない。また、気体のように構造が柔らかすぎたり、ランダムネスが強すぎたりすれば、不安定化して情報を保持できない。さらに、外部からネゲントロピーが降り注ぐとき、いろいろな意味で一様でなく、条件が多様でなければならない。なぜならそういう多様さがなければ、条件の違いによる自然淘汰が起きないからだ。
そう考えると、地球環境は奇跡的にそうした条件を満たしていた。まず、原始地球には水が存在していた。水の存在は非常に重要である。水は液体として柔らかさを持つ。固体のように固すぎず、気体のように柔らかすぎない。生物の柔らかさはそこから来ている。また、水は電気的な極性を持ち、溶媒としての能力が高い。つまり、水が存在すると化学反応が起きやすいということだ。水の中に多様な物質があることでさまざまな化学反応が起きる。これによって高分子が合成されていく。(そのストーリーはイングランドの散逸適応やRNAワールド仮説の説明にある通り。)
水にはこの他にも、「比熱が高い」、「固体(氷)の方が液体(水)よりも密度が小さい」、「2個のHと1個のOが結びついた単純な構造である」(たくさんのC、O、Hが結びついた有機物より合成されやすい)などの性質がある。さまざまな媒質の中で水は非常に特別な性質を持った物質であり、その存在は生命の誕生や進化に欠かせなかったと思われる。
生命の3要素(進化、代謝、境界)を考えても水の存在は重要で不可欠なように見える。地球に水(海、川、湖)があったから、RNAという情報を載せられる高分子が誕生し、遺伝現象が始まった。地球に水があったからこそ、システムの状態が流動的になり、自然選択による適応が始まった(進化)。地球に水があったからこそ、化学反応が起き、代謝が発生した(代謝)。地球に水があったから、柔らかく流動性を持った膜が生まれた(境界)。
水が生命の誕生にとって不可欠な要素であることは広く受け入れられ、すでに良く知られている。太陽系を含め恒星系を考えたとき、水が存在できる範囲は限られている。それは基本的に恒星からの距離で決まり、恒星に近すぎると蒸発してしまい、恒星から遠すぎると氷になってしまう。水が存在するために適度な距離の範囲は「ハビタブル・ゾーン」と呼ばれている。さまざまな恒星の周辺領域の中で水が(つまり生命が)存在しうる範囲のことである。
「水の存在」という条件を一つとっても、宇宙の中で条件を満たす領域がこれだけ狭くなる。最適化の条件[1]〜[3]が「水の存在」以外にどのような自然条件に対応するのかわからないが、それらを満たさなければ、そこで生命が誕生するのは相当難しいだろう。それが地球以外で生命が見つからない理由だと考えられる。
こうした主張の多くは、すでに専門家が言ってきたことを繰り返しているにすぎない。ワールド仮説にしろ、水が重要ということにしろ、進化や学習がアルゴリズムであることにしろ、どれも本書のオリジナルではない。本書が述べている新しい主張は、(1)アルゴリズム、
特に最適化アルゴリズムを自然現象だと見なそうということであり、そういう見方を採用すれば、
(2)ネゲントロピーを利用しながら宇宙の中で進化や学習という自然現象が発生するメカニズムが見えてくるのではないかということである。
「ネゲントロピー」と「アルゴリズム」。この2つが宇宙における生命と意識の誕生を理解するためのキーワードである。これが本書の見方である。
・転がる石と生命の違い
第2部の冒頭で、歩くタンパク質であるミオシンVの動画を見て衝撃を受けたという話をした。その際、「坂道を転がる石と生命のはたらきに何の違いがあるのか」と自分に疑問を投げかけたことも述べた。その答えは、結局、どういうことになるだろうか。ここまで私なりに考察し、たどり着いた答えを、現時点での私の所在地として述べておきたい。
坂道の石の転がりもミオシンVの二足歩行も、半分は同じである。それは宇宙が平衡状態から遠く離れ、ネゲントロピーで満たされた宇宙だから発生した現象である。どちらもネゲントロピーを消費していく。石は転がりながら位置エネルギーを運動エネルギーに変え、さらに斜面のデコボコや他の石と衝突するなどして運動エネルギーを熱エネルギーに変え、やがて停止する。つまり、ネゲントロピーを使い切ったら終わりの現象である。
ミオシンVの歩行もその現象だけで見たら、同じである。ミオシンVの動画を見た後、その動作原理を調べたが、次の通りであった[**]。まず、動作原理について簡単に説明すると、それには力学的なメカニズムと化学的なメカニズムの両方が関わっている。二足歩行する際に二本の足が(道となっている)アクチン線維と結合したり離れたりするが、それは化学的なメカニズムである。その際、ATPのエネルギーが使われている。
また、足の動きには前方と後方で力学的な非対称性がある。後方では前方に引っ張られるためアクチンからの解離が速くなる。その一方、前方では後方に引っ張られるためアクチンの結合が安定化する。この両足間の力学的な相互作用の協同性が一方向の運動を実現するとのことである。
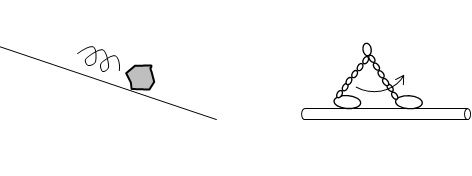
図2_14. 転がる石と二足歩行するミオシンV
残念ながら筆者はまだこの非対称性のしくみを理解できていないのだが、鳥谷部祥一は次のように解説している[**,p.154]。(括弧内は筆者(白井)による。)
「ATP分解の役割はサイクルの方向付けである。ミオシンVは二つの頭部(足のこと)からなるので複雑だが、ATP分解による自由エネルギー変化が方向を決めているのは同じである。…(略)… 自由エネルギーが大きく減少する。これが方向付けをする。」
(鳥谷部祥一、「生物物理学」、日本評論社(2022), p.154)
自由エネルギーという新しい言葉が出てきたが、(一定の条件を満たしていれば)ネゲントロピーとほぼ同じ意味だと考えて良い 。したがって、鳥谷部が言っていることは、ATP分解を通してネゲントロピーが消費されており、それが方向性を決めているということである。
このように坂道を転がる石と二足歩行するミオシンVは、そこだけ見ていれば大して変わらない。ミオシンVはネゲントロピーを消費しながらアクチン線維の下流へと(石が転がるように)二足歩行していく。
それでは、坂道を転がる石と二足歩行するミオシンVは同じようなものだと言ってしまって良いのかと言えば、それはまったく違う。なぜなら坂道を石が転がる現象は、他の現象と無関係な単独の(一回きりで終わりの)現象だが、ミオシンVの二足歩行は、他の現象と関連した一連の生命現象の一部である。わかりやすく言えば、生命という壮大なアルゴリズムの一部なのだ。
「生命と非生命」のセクションで見たように、例えば「代謝」という現象だけ取り出してみれば、生命以外にも多くのシステムに見ることができた。このように、生命現象は一部だけ切り出して見れば、石が転がるだけのような他の現象と同じように見えてしまうことが多い。しかし、重要なことは、その部分的なシステムや現象が、「生命」という大きなアルゴリズムの一部を構成しているという点である。ミオシンVが細胞の中を二足歩行してくれるおかげで、小胞輸送が成り立っている。私が今、指でパソコンのキーボードを叩くとき、指の筋肉が動いてくれるが、それも筋肉細胞の中でミオシン?がアクチン線維の上を歩いてくれるおかげである。
局所的に見ればミオシンの歩行は石の転がりと同じだが、大局的に見ればそれらはまったく違う意味を持つ。ミオシンの歩行は生命という大きな自然アルゴリズムの一部なのだ。それは、生命という自然アルゴリズムの中で、なくてはならない小さなひとかけらである。これが、現時点で私がたどりついた考えである。
========================
【コラム】自然現象としてのアルゴリズム
生命誕生に対していろいろ述べたが、その内容について少し説明をさせてほしい。その理由は、途中からかなり自分のオリジナリティの考え方を入れてしまったからである。それがどこであり、どういう意図だったかを説明しておきたい。
まず、第二章は「2.3 生命の誕生:物質から生命にいたるモデル」までが、普通の生命科学者のさまざまな生命誕生モデルを紹介した。そこには「アルゴリズム」の概念がほとんど入っていない。普通はこう考えますよという紹介だった。次の「2.4 偶然と必然の役割」は、いわゆる複雑系科学者の考え方を紹介した。生命科学者の考え方ほど一般的ではないかもしれないけれど、非常に面白いので紹介した。
問題は、最後の「2.5 第二の飛躍:アルゴリズムの発動」だ。ここには私独自の個性的な考えがたくさん入っている。それを知っておいてほしい。その独自性がどんなものか述べておく。
まず、アルゴリズムの概念を自然現象の一つと見なしているのは本書だけかもしれない。普通、アルゴリズムは計算科学の概念と見なされている。つまり、人間の作業のための概念だ。自然現象ではない。それを、自然現象の一つとして概念を拡張し、生命の誕生にからめて議論したのが本書だ。引用文献を見ても、2.4までは非常に多くの文献が引用されているが、2.5からは少ない。そもそもアルゴリズムを自然現象の一種として位置付けるなど、普通はしないのだ。
しかし、それが必要で重要だと著者の私は考える。生命や学習は自然現象として認められているが、同時に、ある種のアルゴリズムとしても表現できる。したがって、そこにはつながりがあるはずだ。しかし、その議論は非常に少ない。その理由は、アルゴリズムを自然現象の一種と見なすことが、(計算科学以外の)一般的な自然科学においてほぼされていないからではないかと思う。
本書では、アルゴリズムの概念を計算科学の枠組みから飛び出させ、物理学や生物学を含む一般科学の基礎概念として拡張した。自然現象と見なされたそのアルゴリズムを「自然アルゴリズム」と呼び、それが生命の概念の本質の一部だと強く主張した。ここは、2.4までと比べ、独自性の強い主張だから注意してほしい。
ただし、この考えは結論ではない。むしろ新しい種類の議論の出発点として提示したかったのだ。生命科学者たちとは違う見方の一つとして提示したかった。例えば、化学者や生物学者は生命を語るとき、不可逆過程の存在を当たり前のように仮定しながら議論するが、それは当然として良いことだろうか。物理学者も現象を議論するとき、不可逆過程の存在を当たり前のように仮定することが多い。しかし、その概念(不可逆性の概念)が人間という観測者から独立して存在するかと問われれば、物理学者は他分野の科学者たちのように簡単には認めないだろう。観測者としての人間がいるからこそ、不可逆性や時間の流れの概念などが成り立つと考える物理学者は多い。そうした中で生命の起源や生命という不可逆現象を(観測者に依存しない)客観的な自然現象と見なすためには、アルゴリズムの一般化(自然アルゴリズム)がどうしても必要になるというのが私の主張である。
第1章で、時間の非対称性(自然現象の不可逆性)が、観測者による主観性の現れと見るのではなく、宇宙の初期条件と終条件の差から来る本質だと見なすことができると説明した。そこから引き続く第2章で述べたかったことは、その非対称性によって自然現象には必然性(可逆性)と偶然性(不可逆性)の両方が入ってくる。それをもとに、生命(不可逆性を本質として持つ)の誕生について考える場合、偶然性と必然性が理解のカギとなる。
そして、アルゴリズムを自然現象(自然アルゴリズム)として拡張すれば、その偶然性と必然性を次のような形で理解できる。まず、生命の誕生は、時間非対称な宇宙という物理学的な視点から見たら、なかなか起きづらい現象である。宇宙で一般に起きやすいのは、単純なエントロピー増大現象である。生命のように部分系が個体としてエントロピー増大を防ぐような特殊なシステムは誕生しづらい。
そのような誕生しづらい生命が誕生した理由は、自然現象としてのアルゴリズムがはたらく環境がたまたま(偶然)生じる場合があるからである。宇宙には多数の恒星や惑星や衛星が誕生する。これはこの宇宙の必然である。そのさまざまな環境に応じて、いくつかの自然現象が繰り返し発生し、その中で「Aであれば次にBが発生する」というような自然現象のアルゴリズムがしばしば成り立つ。その自然アルゴリズムが発達し、ある水準まで(偶然に)到達すると、そこからそのシステムの発達は必然となる。それが進化だ。これが第2章の内容だった。
========================
第2章の参考文献
(第2章の参考文献は、まだ、調整中です。文献番号と本文のずれや、
引用していない文献タイトルが混ざっていたりするかもしれません。
まだ、作成途中であることをご承知おきください。)
[1] D. クリスチャン(柴田裕之訳)、「オリジン・ストーリー」、筑摩書房(2019)
[2] E. シュレーディンガー(岡小天、鎮目恭夫訳)、「生命とは何か −物理的にみた生細胞−」、岩波文庫(1943)
[3] 佐藤健、「進化には生体膜が必要だった」、裳華房(2018)
[4] 吉川浩満、「理不尽な進化」、ちくま文庫(2021)
[5] H. Naraoka et al., “Soluble organic molecules in samples of the carbonaceous asteroid (162173) Ryugu,”Science, 379, 6634 (2023)
[6] 山岸明彦、高井研、「対論!生命誕生の謎」、集英社(2019)
[7] W. Gilbert,“Origin of life: The RNA world,” Naturem, 319, 618?618(1986)
[8] 田中博、「生命と複雑系」、培風館(2002)
[9] P.ウォード、J.カーシュヴィンク(梶山あゆみ訳)、「生物はなぜ誕生したのか」、河出書房新社(2016)
[10] S. A. カウフマン(水谷淳訳)、「WORLD BEYOND PHYSICS」、森北出版(2020)
[11] Damer and Deamer, “Coupled Phases and Combinatorial Selection in Fluctuating Hydrothermal Pools: A Senario to Guide Experimental Approaches to the Origin of Cellular Life,” Life 5, 1, pp.872-887 (2015)
[12] N.レーン(斉藤隆央訳)、「生命の跳躍」みすず書房(2010)
[13] 長谷川洋、北村成寿、「太陽風を大口で食べ続ける磁気圏」、ISASニュースNo.429(2016)
[14] J. ラブロック(星川淳訳)、「地球生命圏」、工作舎(1984)
[15] S. A. カウフマン(河野至恩訳)、「カウフマン、生命と宇宙を語る」、日本経済新聞社(2002)
[16] P. Lostroh, “Molecular and Cellular Biology of Viruses,” CRC Press (2019)
[17] 田中博、「生命 進化する分子ネットワーク」、パーソナルメディア(2007)
[18] 松岡由希子、「世界初、自己複製する生体ロボット、カエルの幹細胞から開発される」、ニューズウィーク日本版2021年12月1日(2021)
[19] 元田光一、「ロボットが子孫を残すようにコピーを作っていく自己増殖型ロボットの可能性」、未来コトハジメ2020.10.27(日経BPウェブメディア)(2020)
[20] 日経サイエンス2010年10月号SCOPE & ADVANCE、「人工細菌は生命を知る道具」、日経サイエンス社(2010)
[21] 藤崎慎吾、「我々は生命を創れるのか 合成生物学が生み出しつつあるもの」、講談社ブルーバックス(2019)
[22] 資源エネルギー庁、「太陽とCO2で化学品をつくる『人工光合成』、今どこまで進んでいる?」、資源エネルギー庁ホームページ、スペシャルコンテンツ2021-03-04(2021)
[23] 大谷紀子、「進化計算アルゴリズム入門 生物の行動科学から導く最適解」、オーム社(2018)
[24] 小林徹也、杉山友規、「増殖進化適応系に内在する情報熱力学的構造」、生物物理,57(6),287-290(2017)
[25] J.モノー(渡辺格、村上光彦訳)、「偶然と必然」、みすず書房(1972)
[26] N.レーン(斉藤隆央訳)、「生命、エネルギー、進化」、みすず書房(2010)
[27] H. リーヴズ(宇田川博訳)、「宇宙・エントロピー・組織化」、国文社(1992)
[28] 小林武彦、「生物はなぜ死ぬのか」、講談社(2021)
[29] ブルーバックス編集部、「世界最古の生命は、どんな痕跡を残しているのか」、ブルーバックス記事一覧2022.07.14(2022)
[30] G. E. Marsh, ``Thermodynamics and the Origin of Life," Canadian J. Phys., 100, 6 (2022)
[31] I. プリコジン、「存在から発展へ」、みすず書房(1984)
[32] J. L. England, ``Dissipative Adaptation in Driven Self-Assembly,” Nat. Nanotechnol, 10, 919-923 (2015)
[33] J. L. England, ``Statistical Physics of Self-Replicaiton,” J. Chem. Phys., 139: 121923 (2013)
[34] D. Kondepudi and I. Prigogine,“Modern Thermodynamics: From Heat Engines to Dissipative Structures,”John Wiley & Sons, UK (2015)
---------------------------------------------------------