2.4 偶然と必然の役割
・進化は偶然か必然か
物質から生命への変化がどのように起こったか、研究者たちのモデルを見てきたが、ここで生命の誕生や進化が偶然だったのか、必然だったのかについて議論しよう。
まず、進化から考えてみる。進化は「偶然」に支配された現象だろうか。それとも「必然」に支配されているだろうか。「進化は最適化アルゴリズムの一種」と上で述べたが、実際、「進化」を模倣した「遺伝的アルゴリズム」というプログラムがあり、それによって問題が解けることが知られている。なぜ進化を模倣したアルゴリズムで問題が解けるのかというと、突然変異という偶然と自然選択という必然が入っているからである。突然変異は遺伝子コードに偶発的な変化をもたらす。一つのコードから始まったサンプル群は、世代が進むにつれて情報空間の中で拡散し、解を探索する。情報空間には適応度が与えられていて自然選択が起きる。より適応度の高いコード(より良い解)のサンプル数は増え、適応度の低いコードは減る。そのため、最後に一番サンプル数の多いコードを選べば、それが最も良いコード(最適解)となる。こうして進化アルゴリズムを用いて問題を解くことができる。計算は毎回、一つの解にたどり着くのだから、進化アルゴリズムは必然が支配していると言えよう。
進化のダイナミクスを分析するため、さまざまな種類の進化シミュレーションが行われているが、そこでは進化モデルに応じて偶然と必然の両方の効果によって集団(種)の分布が変化していく[20]。最新の研究では、進化適応過程が情報熱力学との類似構造を持つことが指摘されている[21]。これが本当なら、進化は何らかの熱力学的な必然性に支配されていることになる。
現実の地球上での進化はどうだろうか。この疑問に対し、レーンは次のように述べている[23]。
「進化の分子的メカニズムと地球の生命史についてはかなりよくわかっているが、その歴史のどの部分が偶然−ほかの惑星ではかなり違った軌跡を辿ったかもしれない−で、どの部分が物理学の法則や制約に決定づけられているのかは、ほとんどわかっていないのだ。」
(ニック・レーン(斉藤隆央訳)、「生命、エネルギー、進化」、みすず書房(2016)、p.23)
物理学的に考えた場合、次の意味で現実の進化は「偶然」と「必然」の両方に支配されていると言えよう。まず、「偶然」の要素として、彗星の衝突、破滅的な病原体の発生、火山の大爆発など、偶発的な出来事が考えられる。現実の生命の進化にはそれらが大きな影響を及ぼしながら現在の生物種に至っているので、もう一度同じ原始地球を用意し、進化を再現したとしても、それらの影響で同じ進化の経路をたどることはないだろう。
カオスも考慮すべき要素である。カオスとは、初期値のわずかの違いが、大きな結果の違いを生み出す現象である。理論は決定論(=必然)でも結果は非決定論(=偶然)という点が面白い。1993年の映画「ジュラシックパーク」にカオス理論の数学者が登場し、手の甲に落とした水滴が予測不可能なことを使ってカオスが何かを説明する。カオスを進化に適用すると、たとえほんのわずかでも地球の初期条件に違いがあれば、現在の生物種に大きな違いが生まれることになる。
それでは、現実の進化には「必然」の要素がないのかというと、それも間違いだろう。必然も存在する。例えば、オーストラリア大陸での有袋類の進化が、他の大陸での哺乳類の進化に似ていることからわかる。進化における必然性は「ニッチ」や「適応度地形」という形で遺伝子の情報空間に存在し、物理学で言うところの「エントロピー最大」や「自由エネルギー最小」と同様の原理に基づいて、情報空間の中を酔歩しながら最適解へと近づいていくのである。つまり、生物種の分布(生態系)は全体としてある決まったところへと導かれていく。
コンピューター上の仮想空間と現実の空間には大きな違いがある。それは、地球上では彗星衝突や火山爆発など宇宙や地球内部の影響を受けるが、進化シミュレーションはそれらを含まない点である。

図2_8. 現実の世界では偶然、彗星の衝突や火山の大噴火が起きる。
彗星の衝突にしろ火山の爆発にしろ、「偶然」の起源はすべて宇宙の非平衡性にある
という点をここで指摘しておきたい(図4-5)。リーブズ[24]は、平衡から外れたとき、未知の性格が顔を出すと言った。この「未知の性格」が「偶然」である。つまり、宇宙で起こるすべての「偶然」の起源は、宇宙の急膨張に伴う過冷却によって宇宙が非平衡状態となり、大量のネゲントロピーが発生したことにある。彗星の衝突にせよ、火山の爆発にせよ、そのそれぞれが、宇宙が平衡状態へ向かう際に起きる「偶然」の一つである。それらはすべて不可逆な現象で、そうした多くの偶然によって宇宙は徐々に平衡状態へと近づいていく。偶然生じる大絶滅も、その後の大繁栄も、それらはすべて、宇宙が平衡からはるかに離れた状態にあることからくる不可逆現象である。
・生命の誕生は偶然か必然か
次に、生命の誕生が偶然か必然かについて考えよう。つまり、もし原始地球と同じ条件の惑星があったら、必ずそこに生命は誕生するのだろうか。レーンは、J.L.モノーの著書「偶然と必然」[22]を引き合いに出して、次のように述べている[23]。(「その後」の後ろの括弧内は筆者(白井)による。)
「フランスの生物学者ジャック・モノーは名著『偶然と必然』を著した。この本では、地球上の生命の起源は突拍子もない偶然の出来事であり、われわれは空っぽの空間で孤独な存在なのだと淡々と論じられている。…(中略)…。その後(モノーより後の時代)、正反対の主張もされた。生命は宇宙の化学反応が必然的にもたらした産物なのだ。ほとんどどこにでも、すぐに生まれるのだと。」
(ニック・レーン(斉藤隆央訳)、「生命、エネルギー、進化」、みすず書房(2016)、pp.23-24)
つまり、モノーは生命の誕生を突拍子もない偶然だと言ったが、必然だと言う生物学者もいるということだ。また、小林は生命誕生の偶然性について、次のように表現している[25]。
「生命が地球に誕生する確率を表すのに、こんな例えがあります。「25メートルプールにバラバラに分解した腕時計の部品を沈め、ぐるぐるかき混ぜたら自然に腕時計が完成し、しかも動き出す確率に等しい。」
(小林武彦、「生物はなぜ死ぬのか」、講談社(2021)、p.40)
さらに、高井は次のように述べている[7]。
「すべての材料を、地球のありとあらゆる環境から揃えることができたなんて偶然は、ありえないと思っています。」
(山岸明彦、高井研、「対論!生命誕生の謎」、集英社(2019)、p.24)
これらの言葉は、なんの仕組みもなしに、ただ偶然だけによって生命が誕生したとしたら、その確率はあまりにも小さいのではないかと生物学者たちが感じていることを表している。生命の誕生には、何か偶然を超えるような仕組みがあったのではないだろうか。
生命の誕生の偶然性について考えるときに頭に入れておくべきことがある。それは、地球誕生から生命誕生までかなり時間が短かったという事実である。地球の誕生が46億年前で、44-40億年前に海が誕生して安定し、生命が誕生したのは36-40億年前頃の海の中だったと言われている(図4-2)[1,26]。これは、海の状態が安定して、わりとすぐに生命が誕生したことを意味する。生命誕生の時期はかなり早いわけだ。そこまでの時間が短いことは「確率が大きい」ことを示唆する。生命誕生の確率が本当に小さいのであれば、なぜこれほど早く生命は誕生できたのだろうか。
筆者はこの問いに3つの答えが思い浮かぶ。第一は「ただの偶然」という答えである。確率は小さいが0でないから、たまたまそれが実現しただけという考え方である。あまりに都合の良い偶然だと批判されるかもしれないが、絶対に不可能な答えというわけではない。
第二は、生命が宇宙から来たとする答え(宇宙起源説)である。私自身は信じがたいのだが、細胞の形をした生命が宇宙空間を漂っていて、それが原始の地球に舞い降りたのであれば、生命誕生までの時間が短いことを説明できる。ただ、これが正しいとすると、なぜ他の天体でも生命が生まれないのかと疑問が生じる。宇宙空間ではアミノ酸など有機化合物が見つかっているが、そこから細胞まではかなり遠い。この説明では、地球以外で生命が見つかっていないという事実と合わない。
しかし、カーシュヴィンク(2016)は、惑星間パンスペルミア説(火星誕生飛来説)を信じている[10,pp.73-77]。彼がそう考える理由は、地球物理学的な知見から、44億年前の地球の海が今よりかなり広かったと考えられるからである。原初の地球は水に満ちた世界だった。しかし、それはリボースの形成にもタンパク質や核酸の合成にも不向きだった。彼の考えによれば、生命は地球で生まれたのではない。まず原初の火星で生命が誕生し、それが地球に飛来し、現在の地球生命へと発展した。火星はその後、生命の住める環境でなくなり、生命のいない火星となった。カーシュヴィンクによれば、10年間の徹底した実験の結果、隕石が火星の表面から地球へ移動しても、熱で殺菌されることなく生命を運びうることが明確に示されているとのことである。
地球での生命誕生の早さに対する第三の答えは、「何か物理的な仕組みがあった」というものである。これが最も説得力のある答えだと筆者は感じる。多くの科学者がこの考え方に取り組み、そのメカニズムを見出そうと努力している。そのいくつかの成果について見ていこう。
・非平衡熱力学:散逸構造と散逸適応
生命の誕生に偶然を超える物理的な仕組みが何かあったとすると、それはどのような仕組みだろうか。そもそも生命が誕生する以前の物理的・化学的過程(前生命過程)とはどのようなものだったのだろうか。前生命過程については3.2節で「ワールド仮説」としていくつか紹介したが、それらは進化生物学者が考えた前生命過程である。ここで紹介したいのは、物理学者が考えた前生命過程である。
生物学者のワールド仮説は「こんな条件になっていたらこういうメカニズムで生命が自然発生できるんじゃない?」というモデルだが、物理学者はもっと抽象化・一般化して考える。つまり、非平衡状態にあるさまざまな系を仮定し、いろいろ条件を変えて計算機でシミュレーションして、普通の物質からどうやって複雑なシステムが自己組織化され、それがどうやって生命になっていくのかを考えるのだ。そういう物理学者の見方の出発点は「非平衡熱力学」にある。シュレーディンガーからI.プリコジンを経て、生命の起源に関する現代の理論へと続いているので、その発展を振り返ろう[27]。
シュレーディンガーは「生命とは何か」(1943)において「生物は負のエントロピーを食べて生きている」[2,p.129]と言ったことはすでに紹介したが、この言葉は生命が非平衡開放系であることを表している。生命は質の良いエネルギー(高ネゲントロピー)を取り入れ、質の悪いエネルギー(低ネゲントロピー)を放出することで自分の体が劣化するのを防ぎ、システムを維持している。
I.プリコジン(1945)は非平衡開放系の物理学を発展させた著名な物理学である。外部環境に依存したシステムについて議論し、「与えられた境界条件が平衡状態への到達を妨げる場合、系は最小散逸の状態に落ち着く」という「エントロピー生成最小の定理」を示した[28,p.97]。しかし、これは平衡に近い非平衡状態に対して適用すべきもので、平衡から遠く離れた非平衡状態には適用できないものだった。
プリコジンは、平衡からかけ離れた非平衡状態にあるシステムでは、エントロピー生成最小から予測されるふるまいとは逆の場合さえあると指摘した。そして、非平衡(エネルギー散逸)がその秩序の源になりうる例をいくつか示した。1970年代にP.グランスドルフとI.プリコジンは、非平衡熱力学を非線形領域へ拡張し、平衡から遠く離れた散逸系に安定状態が存在することを示した。「散逸系」とは(摩擦など)エネルギー散逸のあるシステムのことで、簡単に言えば、ネゲントロピーの消費のあるシステムのことである。散逸系に現れる安定した構造は、外力がシステムを非線形領域に導くときに平衡から離れたところに現れ、プリゴジンはそれを「散逸構造」と呼んだ。
散逸構造のわかりやすい例は「ベナール・セル」と呼ばれる渦状の構造である。ベナール・セルはベナール対流によって生じる。ベナール対流は火にかけた鍋の中で起きるような対流であり、重力とエネルギー散逸の効果によってその中に渦状の構造が自己組織化される。それがベナール・セルである。セルとは細胞のことだが、実際、細胞のように境界を持った構造が現れる(図2-9)。
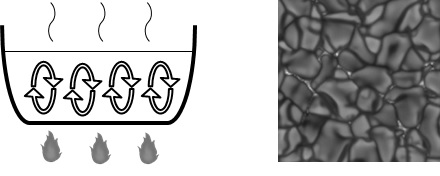
図2_9. (左)ベナール・対流、(右)太陽表面上のベナール・セル(Courtesy of Hinode Scienece Data Centre Europe)
プリコジンは「存在から発展へ」(1984)において自己秩序(安定した構造)の形成の例としてM.アイゲンのモデルを紹介している。そのモデルでは、まず、分子Aと分子Bから作られる高分子が ABABABABAB… という配置を持ち、この高分子を作る反応が自己触媒的であると考える。もし間違って ABAABBAB… という高分子ができた場合、自己触媒によってこの間違いも増殖する。アイゲンは、こうした自己触媒による化学反応の回路網(ハイパーサイクル)が互いに競争することで複雑なものへ進化するのではないかと考えた[27,p.116]。
さまざまな分子からなる非平衡系が、より複雑な高分子の集合へと発展していく場合、そこで成り立つ物理はどのようなものだろうか。その答えとして、J.L.イングランド(2015)は「散逸適応(散逸駆動型適応)」という考え方を提案した[29]。彼は「物理学は“生命的な”性質のタネを播くような形で化学物質を自然に組織化する」と考えた。もしそれが正しければ、(誕生後の進化の道筋は偶然かもしれないが)、生命の誕生は「必然」ということになる。物質から前生物的な化合物が生まれ、それが自己組織化して生命の構成ブロックを作り出し、それが生命へと発展していくのだ。その過程の理解の助けになるのが熱力学ではないかとイングランドは考えた。
彼のアイデアを追ってみよう。今、多数の部品(分子でも良い)が混在するシステムについて考える。このシステムには、熱的な揺らぎだけでなく、システムの変化を「駆動」する揺動的な外力が働いているとする。このとき、系は2つの状態(部品の構成)の間にあるエネルギーの壁(熱揺らぎでは超えられない壁)を外力によって越えて遷移することが可能になる。もし状態が i→j →k と遷移しても、その後、エネルギー散逸がなければ、エネルギーが保存するので k→j→i という逆向きの過程が起きる。しかし、エネルギー散逸がある場合、壁を乗り越える際に使ったエネルギーを熱として放出してしまうので、もとの状態に戻れなくなる。つまり、不可逆になる。
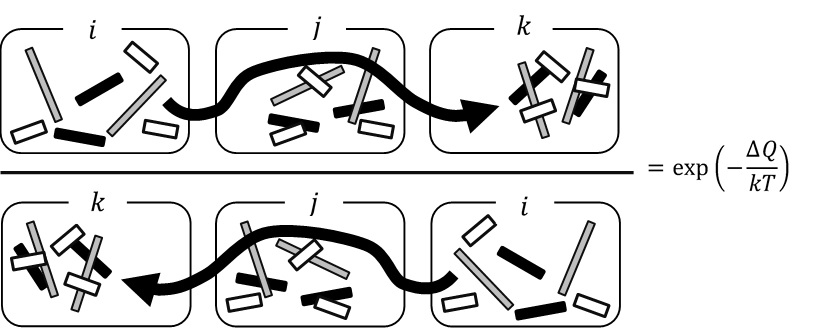
図2_10. 分子集団の状態の順向き経路(上)と逆向き経路(下)の確率
こうしてさまざまな状態変化が一方向的に進んでいくが、どの状態遷移も平等に起きるわけではない。揺動的な外力による仕事をうまく取り入れられる変化もあれば、そうでない変化もある。それは系の構成(分子の構造)に依存する。イングランドによれば、外力からうまくエネルギーを吸収し、散逸できる変化がより成長率が大きくなる。そのため、外力のある環境に最も適応した状態に、構造が自己組織化されていくように見える。イングランドはこれを「散逸適応(散逸駆動型適応)」と呼んだ。
「散逸適応」の概念はまだ十分に検証されたものではないが、生命という複雑なものが現れるための原理の可能性として非常に面白い。この仕組みが本当に自然界に存在するのであれば、「自己組織化への進化は、一般的な非平衡プロセスであり、その最も壮大な現れとして生命の進化がある」というプリコジンたちの言葉がもっともらしく聞こえる[31]。 最近では計算機シミュレーションを含め、さまざまな形で「散逸適応」の検証が始まっている。
・カオスの縁
逸適応の考え方は、どのようにしてばらばらな分子の集合から自発的に複雑な構造が形成され、適応の過程が自然発生するのかを、非平衡熱力学的に基づいて物理学的に説明した。しかし、これだけだと遺伝の要素が説明されていないように見える。システムが自然に自己組織化することや、より適したものが自然選択される(適応)だけでは、進化が始まったとは言えないだろう。やはり遺伝を含む過程がなければ進化とは言いづらい。プリコジンの散逸構造やイングランドの散逸適応系から、どうやって遺伝の過程を含む「生命の進化」が始まったのだろうか。
この難しい問題に対し、複雑系理論やネットワーク理論に基づいた考え方があるので、それを紹介したい。複雑系理論とは、熱現象から生命現象、そして経済現象に至るまで複雑なシステムを共通の枠組みで捉えようとする理論のことである。「複雑系」という名前を筆者が初めて目にしたのは1990年代後半だった。その後、瞬く間に流行となり、2000年代にかなり広がった。実は筆者も複雑系という視点から宇宙を研究し、論文も出版した。
複雑系の研究ではさまざまなものが研究対象となった。例えば、非線形性に注目すれば、太陽黒点の変化から株価のふるまい、水滴の落下などまったく異なる現象を「カオス」という同一の視点から見ることができた。また、多自由度性に注目すれば、ビンからの豆の出方から緊急時の混雑したビルからの避難方法、そして、渋滞現象まで同一の視点から分析することができた。現在は一時期ほどの流行はなくなったように見えるが、分野を横断して自然のしくみを理解しようという試みは今も続いている。本書もその流れを汲む研究の一つと言って良いだろう。
複雑系の研究には、いくつかのテーマがあった。例えば、カオス、ネットワーク、非線形、多自由度、非平衡などである。その研究対象として特に注目を浴びたのが「生命の起源」だった。S.ウルフラムはセル・オートマトンという方法を用いて計算機実験を行い、単純な規則からどのようにして生命と似たような複雑なふるまいが生み出されるのかを調べた。そして、周期的なパターンでもなく、カオスでもないちょうどその間にセル・オートマトンのパラメーターの値があるとき、最も複雑なふるまいをすることがわかった。そして、そこがカオスの境界領域であることから「カオスの縁」と名付けられて、注目を浴びた。
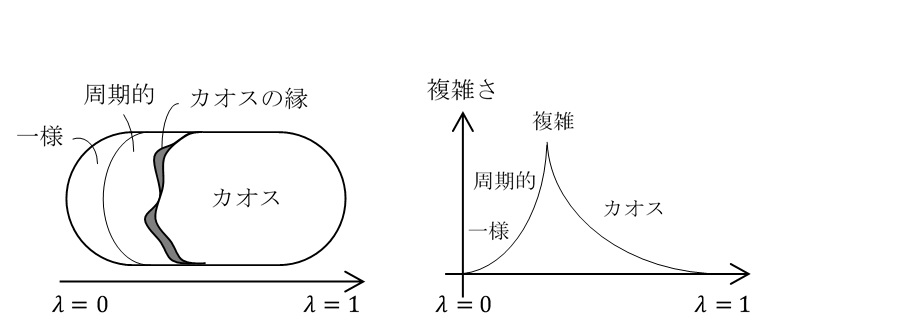
図2_11. カオスの縁のイメージ(横軸はセル・オートマトンのパラメーターの値)
「カオスの縁」辺りでのセル・オートマトンのふるまいはどこか生命に似て、複雑である。その結果を見て、C.G.ラングトンは次のような類推をした[**,p.321]。

この見方が広く受け入れらたかどうわからないが、その後もセル・オートマトンを用いた複雑系の研究は続いているが、「カオスの縁」という概念が必ず生命やその起源の理解に必要なものであるのかどうか、筆者にはわからない。
・自己組織化する複雑ネットワーク
複雑系の研究はネットワーク科学の方向へも発展した。ネットワークはインターネットだけでなく生態系や経済社会、生命個体の内部などさまざまなところで見られ、複雑系の研究対象として興味深いものである。ネットワーク科学の進展により、ネットワークの種類とそこから生じる分布の関係がかなり明らかになった。
生命の起源との関係については、S.A.カウフマンが詳細に考察した。彼はイングランドよりも早く非平衡開放系に着目し、自己構築するシステムの物理学という視点から生命の法則を見出そうとした研究者である。彼は次のように述べている。
「細菌、ゾウリムシ、そして人間のような、環境の中で自分のために行動できるものをここでは「自律体」と呼ぶことにする。…(中略)…、生物圏は誰かが設計し、つくったものではない。この生物圏は、自律体の発生と絶え間ない共進化を通して自律的に構築されたのだ。すべての開かれた熱力学システムに適用できる一般法則がないとしても、熱力学的に開かれた自己構築するシステム―たとえば生物圏のようなもの―に適用できる法則はあるのではないか?」
(S.カウフマン、「カウフマン、生命と宇宙を語る」、日本経済新聞社(2002)、pp.16-17)
彼はその法則を見出す手がかりとして、G.フォン・キードロウスキーの集団的自己触媒集合のモデルとランダム・グラフの理論に着目した。ランダム・グラフの理論は一種のネットワーク理論だ。なぜカウフマンがネットワークの理論に注目したのかは、代謝や自己増殖などの過程を見るとわかる。例えば、ヒトの代謝で起きる化学反応の経路を図に示すと次のようになる。
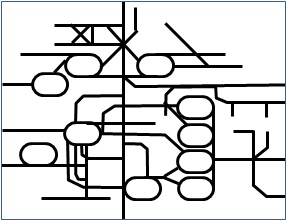
図2_12. ヒトの代謝経路のマップ
これは代謝における化学反応のつながりを表したものだが、複雑なネットワークになっていることがわかると思う。代謝の経路がこのように複雑なネットワークになっているのを見ると、生命の発生を考えるのにネットワーク理論が手がかりになるかもしれないと考えるのは自然なことだろう。
カウフマンはランダム・グラフの理論を調べ、辺と点の比(辺÷点)を増加させていったとき、ある閾値のときに、より大きく連結されたネットワークに相転移することを知った。これを化学反応系のネットワークに当てはめると次のことがわかる。分子A1と分子A2が化学反応を通して高分子Aに変化したとする。この過程をグラフ理論で考えると、化学反応を「点」と見なし、化学反応と各物質を結んだ線を「辺」と見なしたグラフと見ることができる。そして、辺/点の値がある閾値を超えると、化学反応のネットワークが急激に複雑になることが予想される。化学反応のネットワークが相転移するのだ。
カウフマンが参考にしたもう一つの理論はフォン・キードロウスキー集団的自己触媒集合のモデルであり、それは次のようなモデルである。まず、分子A1と分子A2が化学反応して高分子Aが作られる。高分子Aは分子B1と分子B2の化学反応を触媒し、その化学反応から高分子Bが作られる。そして、高分子Bは先ほどの分子A1と分子A2の化学反応を触媒し、再び高分子Aが作られる(図*-*)。こうして互いが互いを触媒しながら自己増殖していくというモデルだ。このような自己触媒反応が集団的に起き、それが生命の起源と関係しているのではないかとフォン・キードロウスキーは考えた。
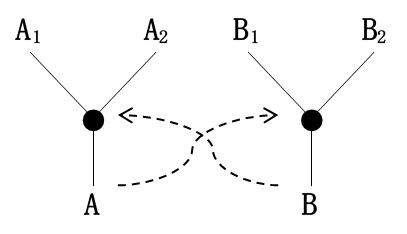
先に述べた「RNAワールド」のように多様な分子が存在する水溶液があったとして、何が起これば生命のような自律体になるのか。そのメカニズムとして、S.カウフマンは集団的自己触媒集合の可能性を考えたのだ。
S.カウフマンはこのような集団的自己触媒集合のネットワークが自己組織化され、それが生命のような自律体の発生の基礎になると考えた。この自律体であれば自己増殖と代謝という生命に不可欠な二つの要素を持っている[2,p.125]。
カウフマンは2020年に「WORLD BEYOND PHYSICS」を出版し、あらためて彼の考えを示した。要点を抽出すると、以下の通りである[**,pp.165-169]。
? 生命は束縛閉回路、仕事サイクル回路、触媒閉回路の3つの回路からなる。生命はこの3つの回路の連動によって自らを構築し、複雑さを高めていく。
? 生命はニッチの生成も自己増殖し、(非エルゴード的宇宙の中で)終わりのない複雑さへと自らを構築していく。
? 奇跡(生命の誕生と進化)を記述する法則は存在しない。生物学を物理学に還元することはできない。
この3つの主張のどこに謎を解く「鍵」が入っているのだろうか。それは?である。そこには、束縛閉回路、仕事サイクル回路、触媒閉回路の3つがあれば自己増殖しながら複雑さを進化するという主張が入っている。
カウフマンの理論を「情報(ネゲントロピー)」という視点から見た場合、どのようなことが言えるだろうか。まず、集団的自己触媒集合のネット―ワークには外部からの「餌」の供給が仮定として含まれている。よって、その「餌」の供給を通して外部からネゲントロピーが供給される。それを利用して自己増幅し、内部の秩序を維持しているのだ。
もう一つ重要な要素である遺伝(進化)についてはどのように説明されているだろうか。カウフマンはデーマーとディーマーのモデル(脂質分子ワールド仮説)に基づいて説明している 。彼らのモデルではウェット=ドライ・サイクルが何百万年も繰り返される中で遺伝システムが生まれる。まず最初に、膜を持つ構造体が多数現れる。そして、乾期と雨期の繰り返しの中でさまざまな分子が切断と結合を繰り返し、徐々にRNAなどの高分子が生まれる。また、乾期には、前生命的システム(膜を持つ構造体)が密集して内容物を共有して生き延びる。そして、雨期には、その内容物が新たな前生命的システムに捕らえられて分裂サイクルが起きる。この過程が一種の自然選択として作用する。何らかの方法で安定性を高めるような内容物を持つシステムが生き残って増殖するため、システムと共に内容物が発展し、より堅牢な原子細胞へと発展していく。これが進化の始まりだと考える。
カウフマンの理論はかなり納得のいくモデルだと感じる。もちろんカウフマンの理論がすべてを説明できているわけではないし、観測などで完全に裏付けられたわけでもないので、まだまだ議論の余地は多い。ただ、ワールド仮説の前段階として、イングランドやカウフマンのように熱力学的な段階から考えるのは重要であるし、個人的に興味深い。生命の起源の問題はこれまで生物学者が中心となって議論してきたが、今後は生物学者だけでなく物理学者も議論に加わって発展していくのかもしれない。楽しみな分野である。
---------------------------------------------------------